Frontal Lobe
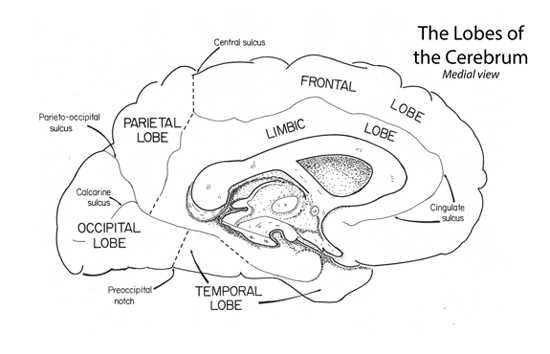
The frontal lobe extends rostrally from the central sulcus to the frontal pole. Medially the frontal lobe extends into the longitudinal fissue to the level of the cingulate gyrys.
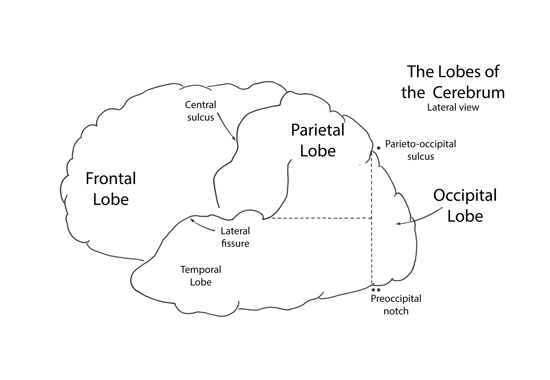
The frontal lobe is composed of numerous gyri. Rostrally on the convexity of the frontal cortex there are three gyri coursing parallel to the longitudinal fissue, termed the superior, middle and inferior frontal gyri. Caudally these three gyri end by abutting the precentral gyrus that courses parallel to the central sulcus (orthogonal to the longitudinal fissure).
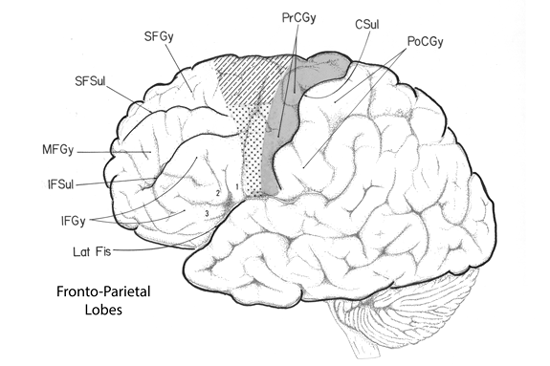
The frontal lobe can also be divided into three functional regions: prefrontal, premotor, and motor cortex. Each region is composed of multiple gyri and sulci. Motor cortex contains the primary motor representation of the cortical hierarchy, while premotor cortex is a secondary level of motor representation in the cerebrum. Finally, the prefrontal cortex is a tertiary or associational level of the cerebrum and is related to executive functions such as decision-making and control of attention.
Motor cortex
Motor cortex is located at the posterior margin of the frontal lobe. It is composed of the primary motor area, situated on the precentral gyrus and the premotor and supplementary motor areas located rostral to the precentral gyrus.
Primary motor cortex
The posterior portion of frontal cortex is the precentral gyrus (area 4) containing primary motor cortex. This region is mainly involved in controlling distal limb musculature and is considered part of the lateral motor system of Kuypers as described in (Nieuwenhuys et al., 1988).
Motor cortex contains the large pyramidal neurons of Betz; it receives afferent fibers from the ventrolateral nucleus of thalamus, the primary somatic sensory cortex, as well as premotor and supplementary motor cortex. It gives rise to corticospinal fibers that terminate in the lateral portions of the ventral horn.
Cells in motor cortex are arranged in a body map; the head is represented lateral and inferior along the precentral gyrus, while the arm is found medial and superior. The representation of the leg is found on the medial aspect of the precentral gyrus as it rolls over the edge of the longitudinal fissure to become the anterior paracentral lobule.
The cells of primary motor cortex exert control over the reflex circuits in the spinal cord as well as directly controlling activity of alpha motoneurons, especially over those innervating the distal musculature of the limbs. Stimulation of primary motor cortex, with weak currents, produces twitches in one or several related muscles in the extremity (Henneman, 1980).
Apparently, one role of primary motor cortex is to encode the force of contraction of the muscle groups about a joint and to prepare these ventral horn neurons to act by adjusting the spinal reflex circuits. In this way, motor cortex is the initiation of the Apower axis@ of the motor system. However, prior to sending instructions from motor cortex to spinal cord, they are tempered with data concerning the programming of muscle groups around surrounding joints, and the guidance of trajectory derived from the supplementary motor and premotor cortices. The programming instructions are derived from the basal ganglia circuits while the trajectory of movement information is derived from the cerebellum. The modified motor cortex instructions are delivered to the spinal neurons to produce smooth, coordinated and balanced movements of the extremities as reviewed in (Kandel et al., 2000).
Premotor Cortex and Supplementary Motor Cortex
The caudal portions of the superior and middle frontal gyri, as well as the rostral border of the precentral gyrus, are involved in motor functions controlling the axial and proximal limb musculature and constitute the premotor and supplementary motor cortex. These two regions are considered part of the medial motor system of Kuypers (Nieuwenhuys et al., 1988). Premotor and supplementary motor cortex are located in area 6 and part of area 8. Afferent projections to these portions of cortex are received from the ipsilateral ventroanterior and ventrolateral thalamic nucleus and from ipsilateral posterior parietal cortex. The premotor and supplementary motor cortices have many targets for their efferent projections. Corticocortical fibers from these two areas interconnect with primary motor cortex bilaterally. Descending corticonuclear fibers from these two areas terminate in the reticular formation of the brainstem, and their corticospinal fibers terminate in the contralateral medial portions of the ventral horn of the spinal cord.
A distinction between premotor and supplementary motor cortex is seen in the subtleties of their afferent connections and their proposed function (Kandel et al., 2000). Premotor cortex receives afferent fibers from the ventrolateral thalamic nucleus (the portion of thalamus receiving the dentothalamic fibers from the cerebellum) and is reported to function by guiding limb trajectory based on sensory information derived from the cerebellum. While the supplementary motor cortex receives thalamic afferent axons from the ventroanterior nucleus (the recipient of pallidothalamic fibers from the basal ganglia) and is reported to function in programming motor sequences derived from the basal ganglia.
Recently, supplementary motor cortex has been divided into two regions: traditional supplementary motor cortex (SMA) and a new region rostral to SMA called presupplementary motor cortex or PreSMA (Luppino et al., 1993). SMA is connected to the motor cortex and projects to the spinal cord; it also demonstrates increased neural activity when we execute a sequence of movements. Conversely, PreSMA primarily projects to premotor cortex and to SMA; it is active during the learning of a movement sequence but not necessarily active during its execution (Hikosaka et al., 1996).
Two specialized areas of motor cortex extend rostrally into premotor cortex. These regions are concerned with eye movements and speech. The frontal eye fields (inferior portion of area 8 plus part of areas 6 and 9) are found on the caudal border of the middle frontal gyrus; they direct the movement of the eyes into the opposite visual hemisphere. On the inferior border of the precentral gyrus and spreading onto the pars opercularis of the inferior frontal gyrus is Broca's speech area (areas 44 and 45;). Broca's area contains the motor patterns for speech. Right‑handed persons have Broca's area only in the left or dominant cortex. Superior to Broca's area is a related region of cortex controlling the hand and containing the motor patterns for writing (see section on aphasias). The hand region of the premotor cortex is termed Exner's hand area.
Premotor cortex and apraxias
In the dominant hemisphere there is a region located at the terminus of the lateral fissure that is involved with comprehension. The is region is often termed Wernicke's area and will be describe in more detail later. The Wernicke’s area receives sensory information from all sensory portions of cerebral cortex. After interpreting these data commands are issued to the motor areas to follow out prescribed movement patterns. Since Wernicke's area is located in the dominant hemisphere, these commands must be transmitted bilaterally. A diagram illustrating a proposed schema of these connections is presented in Figure 12. Wernicke’s area projects to the ipsilateral premotor cortex to control the dominant extremities. However, the control the non-dominant extremities, it projects first to the ipsilateral premotor cortex which then projects to the contralateral premotor region and on to the contralateral motor cortex. Thus instructions concerning volitional control of the non-dominant extremites cross the midline in the anterior portion of the corpus callosum.
It is possible, and indeed relatively common to have a lesion separate Wernicke's area from motor or premotor cortex. When this occurs the patient is capable of comprehending the verbal command, but is unable to instruct the body to carry out the command. Such patients are not paralyzed since they have no demonstrable deficit in their motor system. If the commands require a verbal response and the patient is faulty, they are considered aphasic. However, if the command requires a movement response, such as "raise your right hand" and the patient cannot comply despite their lack of paralysis, they are considered apraxic (Geschwind, 1975).
Given the myriad of command pathways leaving Wernicke's area, there are numerous types of apraxias documented. We will consider only one as an example.
Case Study: A stroke patient with inability to execute verbal commands with the left hand
This is a 41‑year‑old Boston police officer that was admitted to the hospital following a month period of headaches, behavioral changes such as apathy, forgetfulness and confusion. No sensory deficits were detected; motor system was everywhere in tact, no abnormal reflexes, deep tendon reflexes were normal and plantar reflexes were flexor. Pneumoencephalogram revealed a space-occupying lesion in the frontal lobes, craniotomy was performed and a large tumor was removed.
Post‑operative examination of the patient revealed normal power and tone everywhere in the motor system and no profound sensory losses. He could not write with his left hand. When asked to perform a task with his left hand he was unable. Yet he had full range of motion with the limb. He was able to carry out all commands to use the right hand. The patient, who was observed over many months, would carry out commands with his right arm, correctly and without hesitation‑for example, "Show me how you would comb your hair," "Show me how you would use a hammer," "Wave goodby." However, when given commands for his left arm, such as "Show me how you would use a comb with your left hand," he would usually either make no response or would make a clearly incorrect response. He could use his left arm quite normally for these tasks under other circumstances: if the examiner performed the act, the patient could imitate with his left arm, or if the patient were handed a hammer or a comb, he handled them normally.
The fully correct performance by the left arm ruled out the possibility that the failure to respond to verbal command could be a result of weakness, incoordination, or any other elementary disorder involving the left arm. The correct response of the right arm to verbal command ruled out inattention, incomprehension, or uncooperativeness on the part of the patient (Geschwind, 1975).
(case history and quotations taken from (Geschwind and Kaplan, 1962))
Comment on the Case: At autopsy, years later, it was found that the tumor had damages the anterior four/fifths of the corpus callosum, effectively separating the left premotor cortex from the right. The loss of communication between the two hemispheres prevented the premotor area on the left from instructing that on the right side. Consequently, verbal instructions, processed in the left hemisphere, could not reach the right and the left arm remained unresponsive. Mimicry, which uses other cortical routes remained intact and the patient could perform the task while following the examiners lead.
To understand what is happening in this patient we must consider how motor commands are sent to the appropriate cortex. In a normal individual, Wernicke's area receives the auditory information requesting them to wave the left arm. Instructions are sent from Wernicke's, via the arcuate fasciculus to the premotor area of the left hemisphere; from here the information is passed through the corpus callosum to the hand premotor cortex on the right where it is converted into a motor response.
This patient could comprehend the statement "Wave with your left hand, but could not instruct the premotor portion of right cortex to execute the programs necessary to wave with the left hand. The lesion disconnected the premotor region of the right hemisphere (controls left hand) from Wernicke's area in the left hemisphere. Thus although the patient had the motor programs to wave the left hand, and could do so spontaneously, he could not access these programs in response to verbal commands.